|
|||

|
|||
|
Journal ARS Article |
|||
|
Vol. 47: No. 3: Year 1993 |
|||
|
Concerning the Origin and Distribution of Rhododendrons E. Irving and R. Hebda Abstract There are two important features of the distribution of species of the genus Rhododendron that require explanation. First, the overwhelming majority of them occur either on the slopes of the very deep valleys that border the eastern Himalayas and southeastern Tibet, or in the mountain ranges that form the backbones of the archipelago stretching between mainland Asia and Australia - the islands of Java, Sumatra, Borneo, New Guinea, and the Philippines. The second feature is that the remaining species, although far fewer in number, are spread much more widely over the northern hemisphere, occurring in pockets that, to a considerable degree, are isolated from one another - Japan, northwestern North America, the Appalachian, and Caucasus Mountains. We propose that, during their early history, rhododendrons were much more evenly spread than they are now, and that their present discontinuous distribution was caused by the encroachment, in comparatively recent times, of conditions hostile to their existence, namely the extensions of glacial ice and of modern grassland and deserts. We also argue that the present remarkable concentration of species in southeastern Asia has arisen because it is there that habitats were developed in which rhododendrons found not only shelter from climatic vicissitudes, but in which they could flourish and speciate; apparently they were able to do this at a time when rhododendrons elsewhere were being driven from much of their former range. We hope to show how the history of this horticulturally outstanding genus might have been shaped by global and regional geological events. Introduction In this article we set out our ideas concerning the roles that geological, climatic and ecological changes may have played in shaping the present distribution of the genus Rhododendron. It is known that continents move, oceans open and close, and that there are mountain ranges where once the land was flat and low. It is also known that the present climate of Earth is atypical. In the past, climate was generally warmer than now, especially in intermediate and high latitudes. Rhododendrons are a relatively old group of flowering plants and were in existence whilst these vast changes in geography and climate took place. It is likely, therefore, that these changes have been major factors in shaping their present distribution. We shall describe and attempt to evaluate these factors. We confess to having no special knowledge of rhododendrons, neither their taxonomy nor their horticultural history. Much that we have to say is necessarily very speculative. Nevertheless it is our hope that something is to be gained by bringing to bear a geological perspective. Our interest in these problems dates from the time when, at the invitation of Mr. Norman Todd, we began preparing a lecture for the 1989 convention of the American Rhododendron Society in Victoria, and it is at his urging and that of other fellow gardeners that we record here the substance of that lecture. We shall try to grasp the full sweep of the majestic phenomenon of the genus Rhododendron through geological time, to identify the important generalities, and to search for rational explanations of them. The arguments are made in as simple a manner as possible, so that readers will not only be encouraged to persevere to the end, but may even be tempted to determine how well (or poorly) our ideas fit their own knowledge of these remarkable plants. Should readers find our arguments wanting, it is for them to propose more acceptable alternatives. Distribution Rhododendron species circle the globe (Fig. 1). However, the greatest number occur in the pan-shaped area extending along the southern Himalayas east into southwestern China (Fig. 2). This region, which includes parts of Nepal, Bhutan, northeastern India, northeastern Burma, southeast Tibet, western Szechuan and northwestern Yunnan, consists of high steep-sided mountain ranges separated by deep valleys. Nowhere else on Earth are there so many very deep valleys clustered so closely together. We refer to this area as the "region of extreme relief" (RER in Fig. 3(a)). Eastward from this area, in central and eastern China, Korea and Japan, the number of species (that is, the diversity) declines by a factor 10 roughly. Rhododendron species also occur in great numbers in the mountainous backbones of the islands between Asia and Australia, which are referred to as the "high-island archipelago" (Fig. 3(a)). Between the latter and the region of extreme relief, many species occur in the mountain ranges of Thailand, Vietnam and Malaya. Elsewhere, rhododendrons occur in northeastern Asia, in isolated communities in the mountains of southern India, southern Europe, the Caucasus Mountains, northeastern Turkey, eastern, northern and western North America, and across the sub-arctic zone. The number of species is, however, comparatively few.
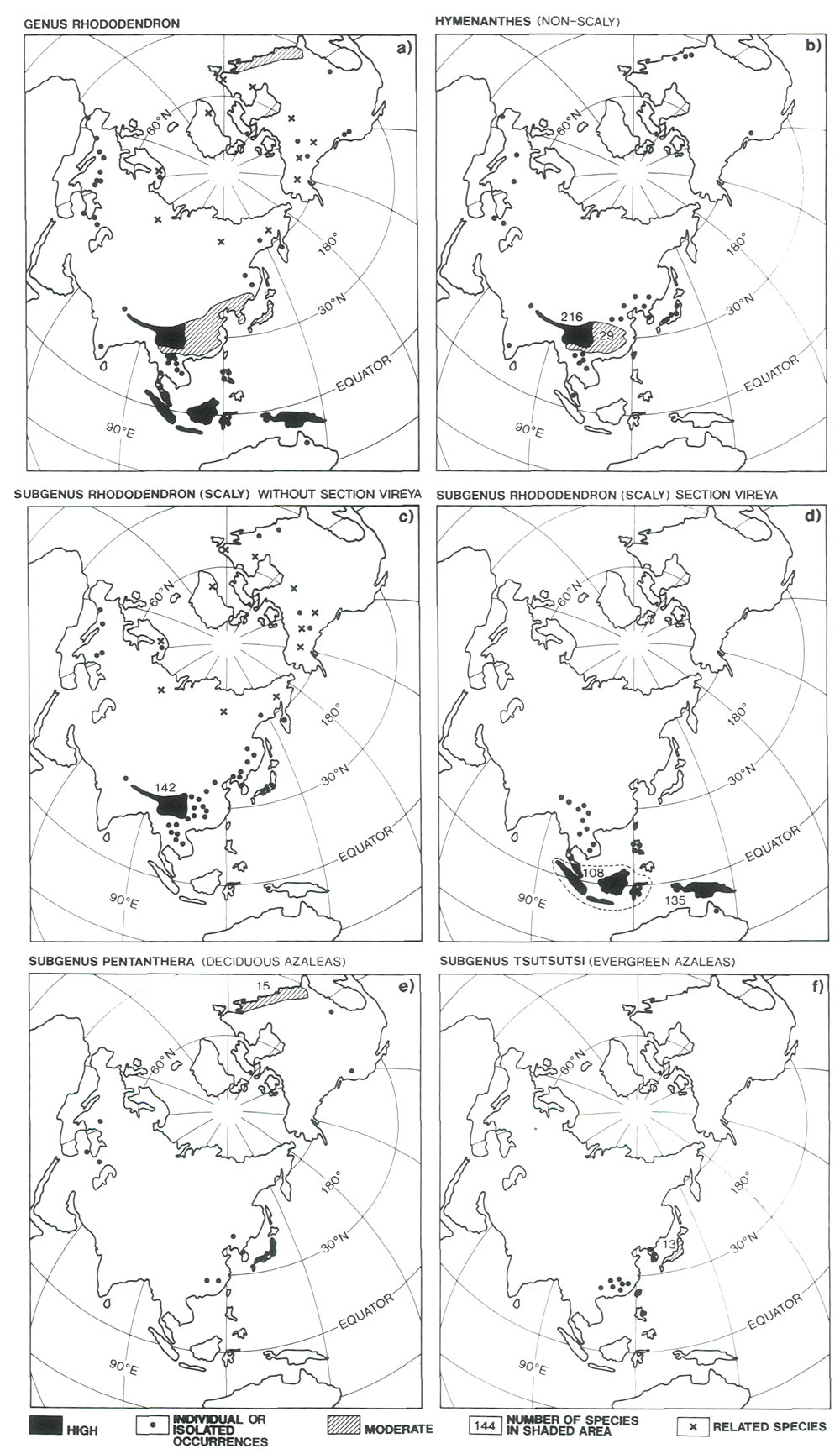
Figure 1. Distribution of species. (a) all rhododendrons; (b) subgenus Hymenanthes, (c) subgenus Rhododendron without section Vireya, (d) section Vireya of the subgenus Rhododendron, (e) subgenus Pentanthera, (f) subgenus Tsutsusi. The compilations (which have been made from Hortus Third (1976), Cullen (1980), Chamberlain (1982) and Luteyn (1980) are not exhaustive, but are considered representative. Cross-hatched regions of moderate diversity have about 10 times fewer species than those of high diversity shown solid black. The dots (isolated species) and crosses (related species) in (c)) do not always denote individual occurrence, but are intended to indicate the wide-spreadness of such species as R. lapponicum, and R. albiflorum, and the related Labrador teas (genus Ledum) which some authors now include in the genus Rhododendron.
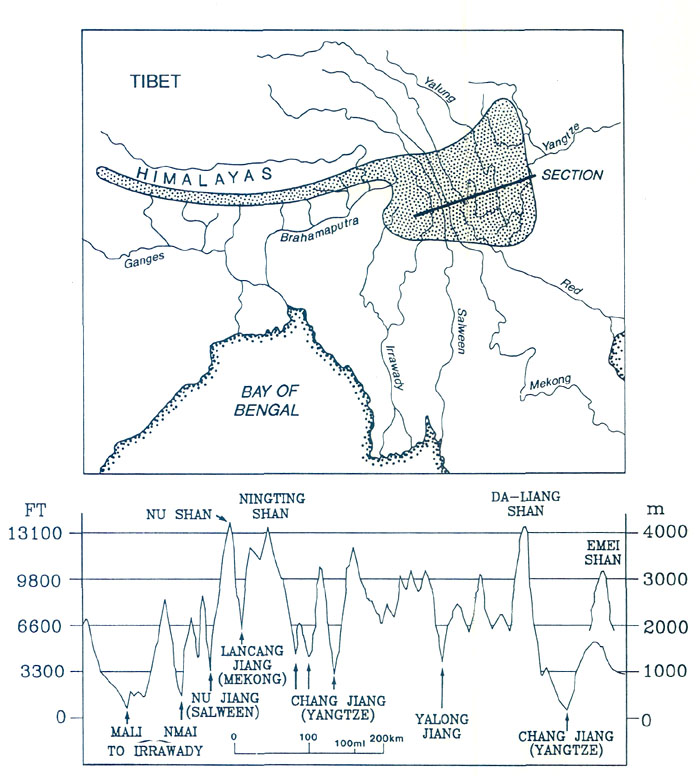
Figure 2. The region of extreme relief and heavy rainfall (stippled in upper diagram). The cross-section below is along the line indicated in the upper diagram. Emei Shan in Szechuan, with its numerous rhododendron species, is a little to the north of the line of section. Eight subgenera of rhododendrons have been recognized, but there are four main ones (Luteyn 1980). Rhododendrons of the subgenus Hymenanthes (elepidote) are evergreen and have large non-scaly leaves. Rhododendrons of the subgenus Rhododendron (Iepidote) have smaller scaly leaves, are usually evergreen, but occasionally are semi-deciduous. The deciduous azaleas constitute the subgenus Pentanthera, and the evergreen azaleas the sub-genus Tsutsusi. Species and hybrids of the subgenera Hymenanthes and Rhododendron comprise what gardeners refer to loosely as "rhododendrons". Those of the subgenera Pentanthera and Tsutsusi comprise the "azaleas". Within each subgenus, rhododendrons generally will hybridize readily. This is especially true of rhododendrons from the region of extreme relief and from the high-island archipelago, indicating that within these populations there are no major genetic barriers and that speciation is currently rapid. Rarely has it been possible to cross rhododendrons from different subgenera. It is probable, therefore, that the four subgenera separated from an ancestral stock early in the evolution of the genus and have long been reproductively separate. It invites confusion to have the word rhododendron denote in one context a genus, in another a subgenus, and in yet another to have it meant in a loose gardening sense, and we shall try always to make clear our meaning. The subgenus Hymenanthes contains about 225 species (Chamberlain 1982). They occur in southern Europe, the Caucasus Mountains, eastern and western North America, but the greatest diversity by far occurs in the region of extreme relief and to a lesser extent in central and south- eastern China (Fig. 1 (b)). The subgenus Rhododendron has about 400 species and is the most widespread of all the subgenera. The section Vireya, which has over 200 species, is found almost entirely in the mountains of Java and Sumatra, Borneo, the Philippine Islands and New Guinea (Sleumer 1972). There is a single representative in Queensland, Australia (Fig. 1 (d)). There is also a string of occurrences in the mountains of Malaysia and Thailand, connecting the high mountain archipelago with the region of extreme relief, which itself supports several Vireya species. Aside from the vireyas, most lepidote rhododendrons occur in the region of extreme relief, with isolated species scattered across eastern and southeastern Asia, southern Europe, northwestern and eastern North America (Fig. l (c)). An interesting member of the subgenus Rhododendron is Rhododendron lapponicum, which has a circum-polar distribution extending from Lapland across Siberia to northern Canada and into the northern USA. An unusual species, R. albiflorum, sometimes placed in its own subgenus, is resident only in northwestern North America, mainly British Columbia. There are also the Labrador teas (Ledum groenlandicum and L. glandulosum), which are closely related to rhododendrons and, according to Kron and Judd (1990), should be included in the genus. The two subgenera of azaleas have comparatively few species and, so far as we are able to determine, are absent from the region of extreme relief. Species of the subgenus Pentanthera (deciduous azaleas) are most numerous in the southeastern USA and Japan (Fig. 1 (e)), with notable isolated species in western USA (the fragrant western azalea, R. occidentale) and Asia Minor (the fragrant R. luteum). Species of Tsutsusi (evergreen azaleas) occur only in southeastern Asia. They are most numerous in Japan, with some species in southeastern China, Korea, the Philippines and in Taiwan (Fig. 1 (f)).
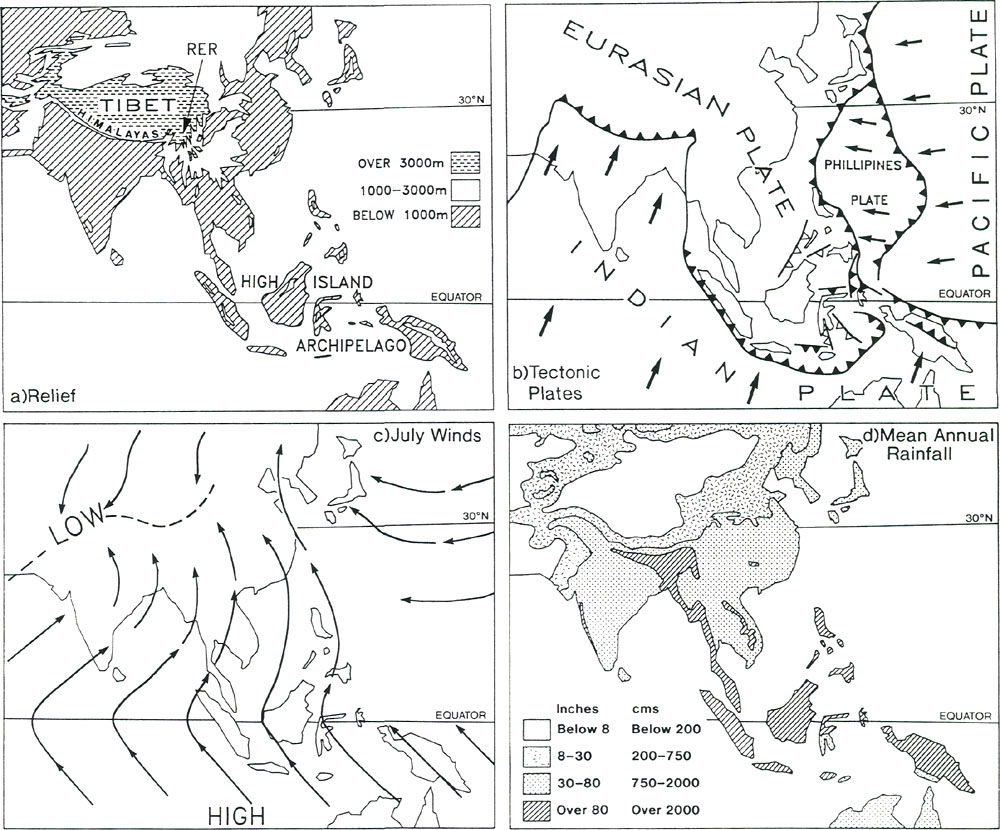
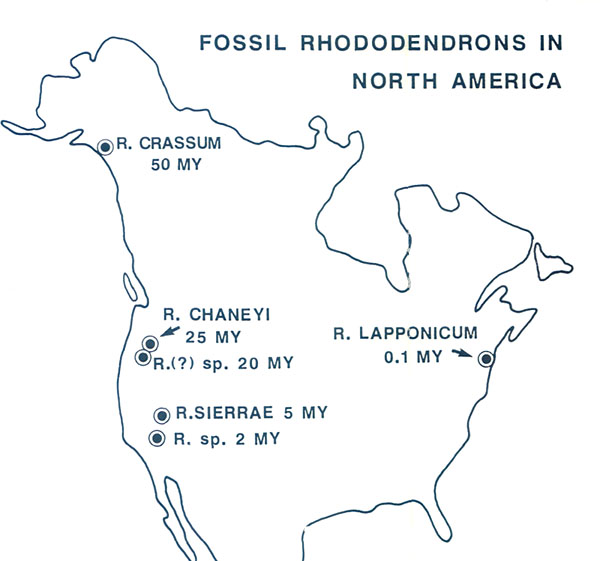
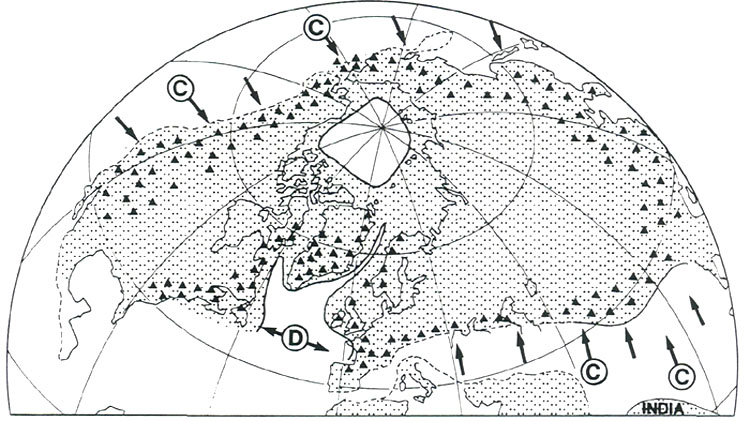
Figure 3. a) Relief of southeast Asia; RER is the region of extreme relief. b) Tectonic plates; bold lines denote plate margins; arrows denote direction of motion relative to the Eurasian plate which is assumed fixed; solid flecks mark the places (subduction zones) where one plate dives beneath another, the overriding zone being on the flecked side; open flecks denote subduction zones that are now inactive but which have been active during the past few million years; plates on either side of margins (transform margins) without flecks are undergoing mainly shearing motion with little or no subsection. c) July winds; high and low pressure regions are marked. d) Mean annual rainfall. Fossils and Habit Rhododendrons belong to the family Ericaceae. There are very few rhododendron fossils. According to Muller (1980) the ericaceous plants (recognizable by their fossilized pollen grains that are arranged in groups of four) appeared first in the Maastrichtian Stage about 68 million years ago. The Maastrichtian is the last stage of the Cretaceous Period and immediately precedes the extinction of dinosaurs. The earliest rhododendron fossils are leaf imprints in rocks of early Tertiary age (about 50 million years old) from Alaska (Hollick and Smith 1944). Other fossils are known from the Tertiary and Pleistocene deposits of North America (Fig. 4). Provided that these fossils have been correctly identified, they establish that rhododendrons have been in existence for at least 50 million years. They could have been in existence before that time, but not before 68 million years ago when the Ericaceae first appeared. This is of interest because it indicates that rhododendrons probably did not originate in the region of extreme relief or in the high-island archipelago (where they are now most abundant and most diverse) because these regions did not exist 50 million years ago (see below).
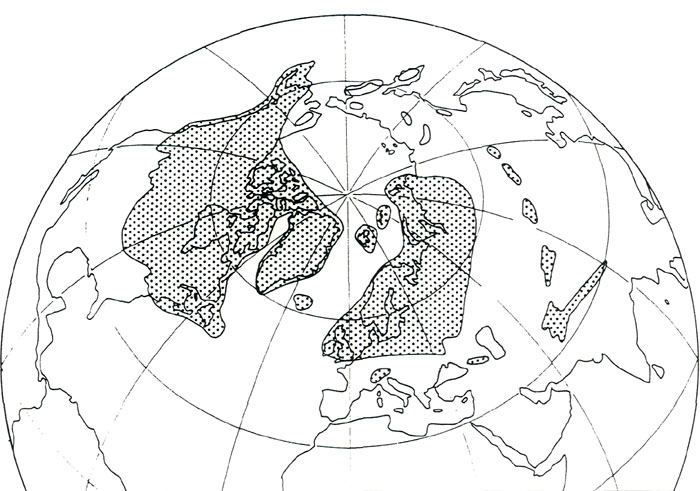
Figure 4. Fossil rhododendrons in North America. Fossil records of Ledum are not included. Compilation from various sources. Nowadays, rhododendrons grow mainly in temperate climates and in moist but well-drained sites. They are not common at low altitudes in hot equatorial forests, although they do occur (e.g., R. brookeanum of Borneo). They are abundant in the mountains of the equatorial high- island archipelago. They are rare or absent in deserts, grasslands, boreal forests or tundra. Except for R. lapponicum (and the Labrador teas if these are regarded as rhododendrons) which has adapted to the climatic extremes of arctic and alpine regions, rhododendrons flourish best in the habitats described above. We shall assume that this has, by-and-large, been so since the genus originated. The scarcity of the fossil record is consistent with an upland well-drained habitat. There is only a slim chance that the remains of modern species will ever find their way into sediments at the bottom of upland lakes, which themselves are vulnerable to erosion and unlikely to become part of the geological record. Hence, it would be expected that fossils would be sparse if rhododendrons had always preferred upland habitats just as they do today. Had they been common lowland plants, they are likely to have been far more prominent in the fossil record than they are. Given that there are so few fossils, it may seem impossible to determine the origins and past distribution of rhododendrons. However, by determining the past distribution of those plant assemblages which grew under conditions amicable to rhododendrons, it is possible to identify those regions of the world where rhododendrons could have flourished during their long history. Today, the temperate, moist, upland settings generally preferred by rhododendrons are found in deciduous or broad-leaved evergreen forest, often with a coniferous component. We shall attempt, therefore, to infer where such upland forests might have occurred during the time that rhododendrons are considered to have existed, that is, during the past 50 million years or so. Continental Drift and the Rise of Mountains Three major physical factors affect the distribution of vegetation through time - global climate change, the drift of continents, and the rise of mountains. The climate of the entire earth can change and affect an individual location, and the climate of an individual locality can change either as it moves from low to high latitude, or as it is uplifted from low to high elevations. These factors are now considered. About 60 million years ago, South America, Africa (which then included Arabia), Antarctica, Australia and India were huge islands, not yet joined to the northern continents (Fig. 5). The Atlantic Ocean was much narrower than now, and its northern part had just begun to open. The gulfs separating Greenland, North America, and Europe were narrow and probably did little to impede the free exchange of land plants. Between Alaska and Siberia there was a continuous land-bridge called Beringia.
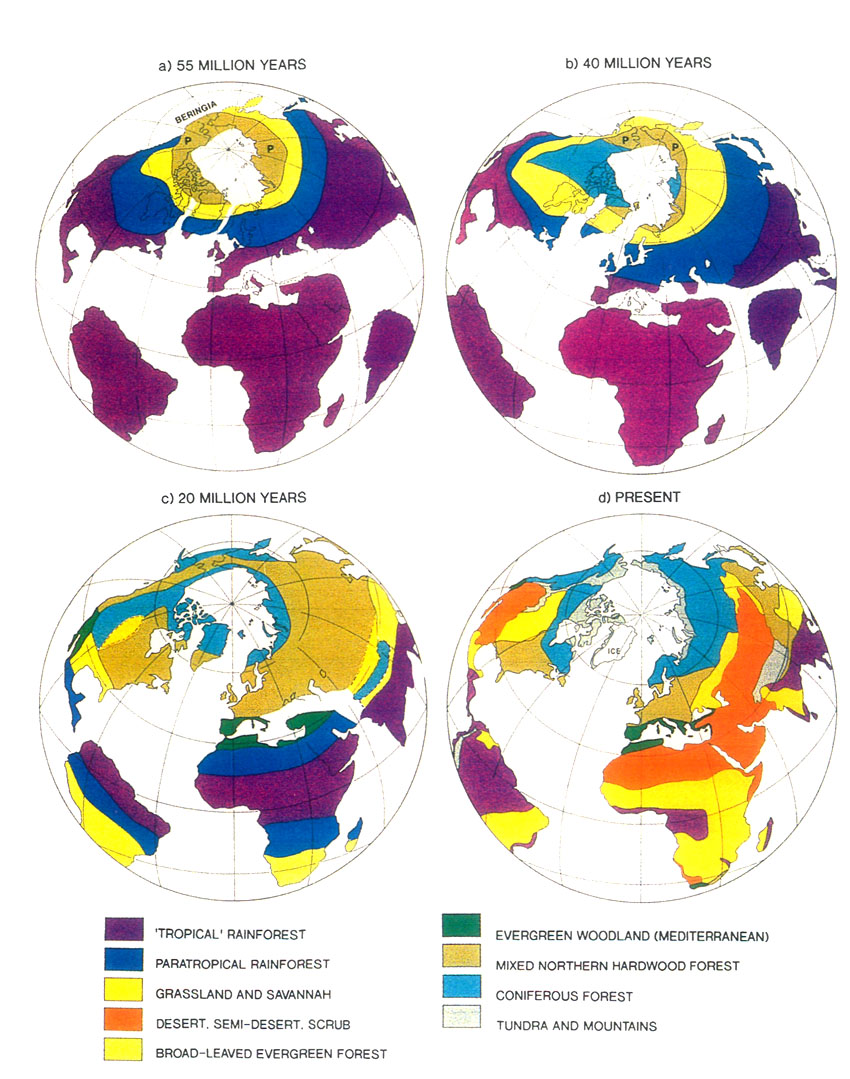
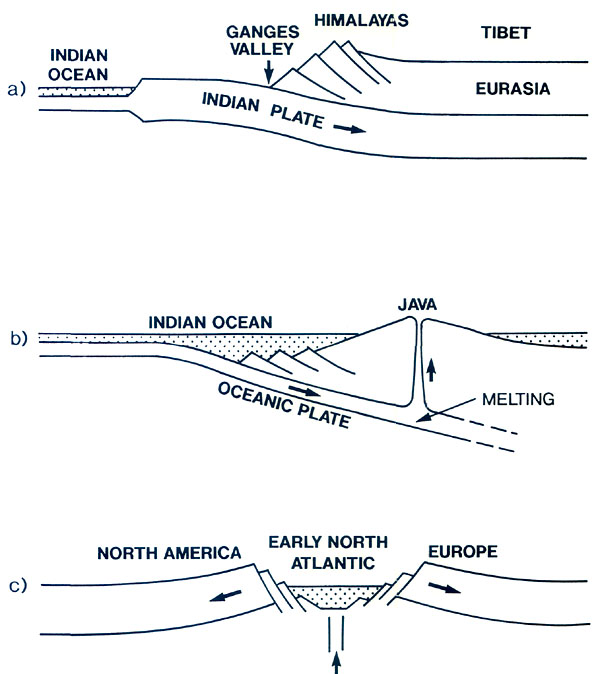
Figure 5 (a). Positions of continents and their vegetational zones. (a) about 60 million years ago, (b) about 40 million years ago, (c) about 20 million years ago, (d) present vegetational zones. Vegetational zones in (a) (b) and (c) have been compiled with simplifications and modifications from Wolfe (1985), and (d) from The Times Atlas. Base maps are from Irving (1982). The old polar deciduous forest has been labeled P to distinguish it from later, botanically different, mixed northern hardwood forests. Upland terrain seems to be an essential ingredient of rhododendron habitats, and therefore something needs to be said about how uplands are created. There are three main processes, the first two occurring when the lithospheric plates of Earth converge, the third when they move apart. The highest mountains are formed when plates carrying continents collide and the lithosphere in the collision zone thickens. Opinions differ as to how thickening takes place, one possibility being that one plate slides (subducts) beneath the other. Continents are relatively light and buoyant so that thickened continent-bearing Iithosphere floats upward, by the principle of Archimedes, creating very high ground (Fig. 6(a)). The second process occurs by subduction of a relatively dense oceanic plate, that is, a Iithospheric plate that does not carry a continent. When such a plate reaches a depth of about 100 km, it melts, producing hot lava which rises and creates chains of volcanoes. The rising lava brings with it much heat which causes expansion, a decrease in density, and, by Archimedes' principle, uplift. Such volcanoes can form islands rising from deep ocean (e.g., Java, Fig. 6(b)), or they can be superimposed on the edge of continents (e.g., Mt. St. Helens). Thirdly, mountains form when a plate breaks and the fragments move apart. The zone of fracture or rift allows molten material to well up, bringing heat from deep within the Earth, causing expansion, lowering density and buoying the surface upward (Fig. 6(c)).
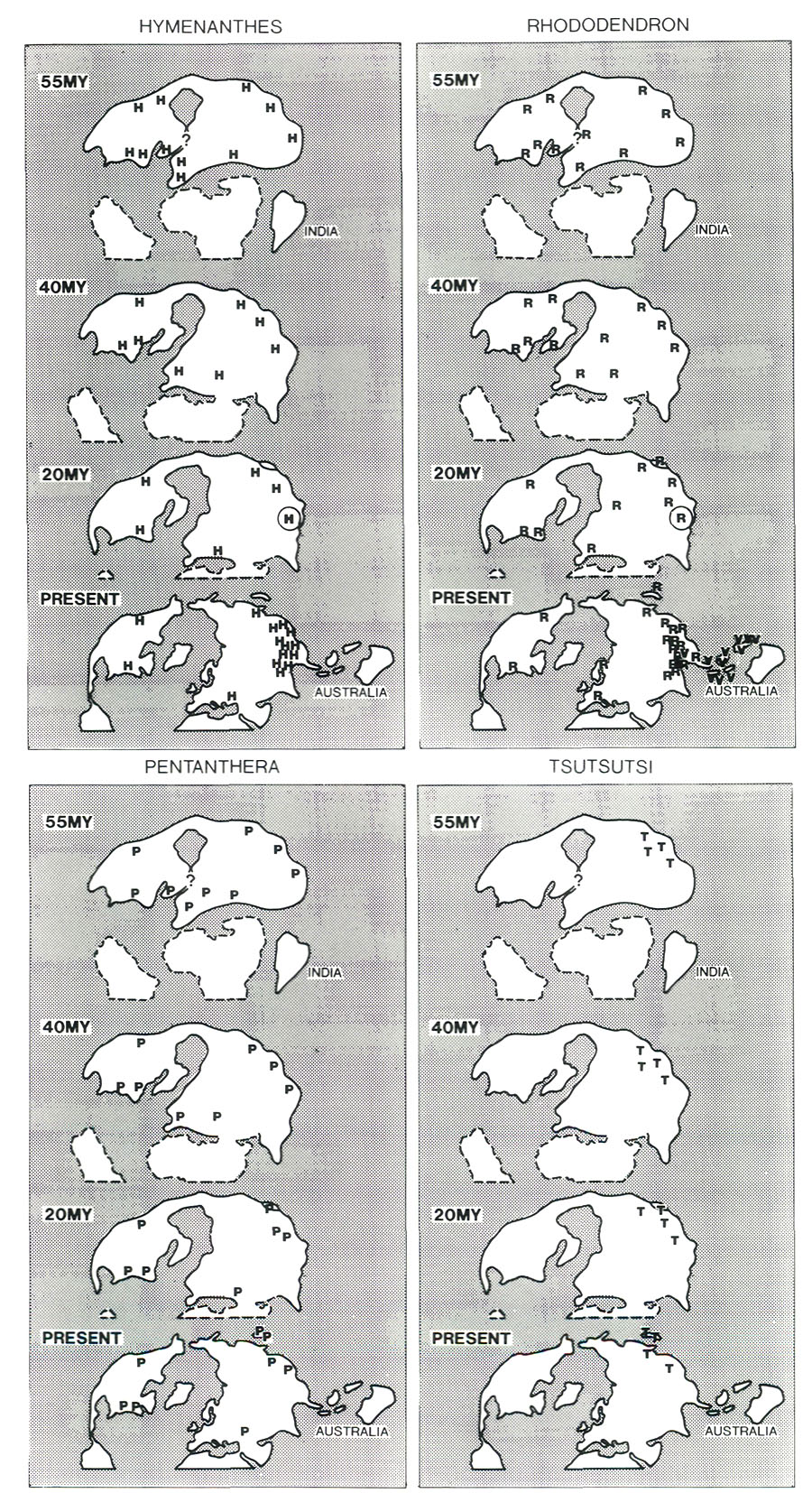
Figure 6. Some ways of forming mountains. The diagrams are much simplified and are intended only to illustrate general principles. (a) Collision of continents; cross-section through India and Tibet showing one possible way of explaining the doubling of the crust and the elevation of Tibet. (b) Subduction of ocean plate and creation of mountainous volcanic arcs or islands; the example shown is from the high-island archipelago. (c) Rifting apart of a continent; cross-section of an early stage of formation of the North Atlantic. Applying these ideas to our knowledge of plate motions and continental positions, it is possible to infer where uplands probably existed during the early stages of rhododendron development 50 or 60 million years ago (Fig. 7). A zone of convergence occurred along the southern edge of Europe and Asia; Africa and India were moving northward but had not yet collided with the northern land mass. A zone of convergence also existed where the Pacific Plate plunged beneath the western border of North America and the east coast of Asia. These zones of convergence probably were marked by huge volcanic chains of the type shown in Figure 6(b). In the south and west, Europe and North America had recently split apart forming the small but expanding North Atlantic Ocean, with gulfs extending northward between North America and Greenland, and between Greenland and Scandinavia. As shown in Figure 6(c), these rifts probably had uplifted mountainous edges. Consequently, it seems likely that the huge semi-continuous land mass of North America and Eurasia (often called Laurasia) was rimmed more or less entirely by broad upland zones, along which rhododendrons during the early part of their evolution would have been free to disperse. By 40 million years ago there was still a land connection between Alaska and Asia (Fig. 5(b)). The Atlantic continued to widen and began to form a barrier to the free migration of land plants. Africa was closer to Europe, and India began to impinge on Asia. Mountain and upland probably continued to have a comparable distribution to that at about 50 million years (Fig. 7). It is important to note that today's major centers of diversity, the region of extreme relief and the high-island archipelago, did not yet exist.
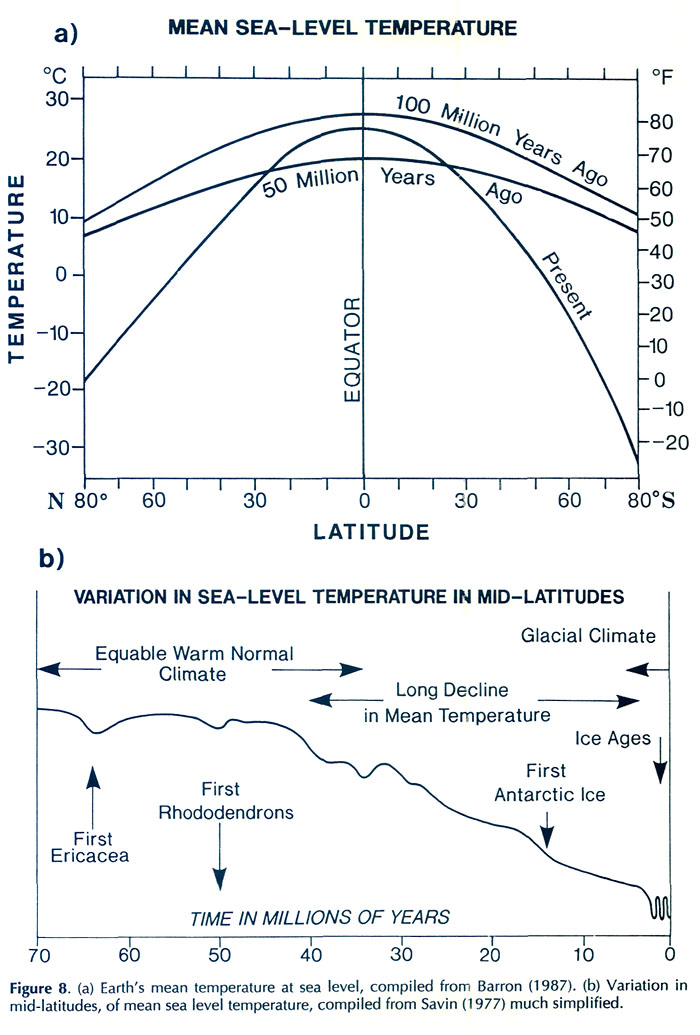
Figure 7. Possible distribution of uplands about 55 million years ago. Arrows denote convergent (C) and divergent (D) margins corresponding to Figures 6(b) and (c). Twenty million years ago, the North Atlantic widened sufficiently to become a barrier to the spread of plants, but the land bridge of Beringia remained (Fig. 5(c)). Most importantly, India began to under thrust Asia, and the processes which, as will be explained shortly, lead to the creation of the region of extreme relief and the high-island archipelago were set in motion. Present geography has been created by continuation of the above processes - the Atlantic continued to widen, South America approached North America, Africa, India, Asia and Europe coalesced to form a more-or-less continuous land mass, and Australia moved up from the south. Global Climatic Change Of course, it is not sufficient to say that because there is a suitably hilly landscape, such as that depicted in Figure 7, that rhododendrons would actually have grown there. A suitable climate is needed also, and the Earth's climate is known to have changed drastically since rhododendrons first appeared. Today there is a year-round ice cap at both poles, the difference in mean surface temperature between pole and equator is over 40°C (Fig. 8(a)), and at latitudes above about 55° the mean surface temperature is below freezing. Earth is said to be in a "glacial period". Earth has been in a glacial period for the past 2 to 3 million years, during which there have been large oscillations roughly every 50,000 years. At times (called ice ages) glaciers have extended down into intermediate latitude (Fig. 9). At other times (called interglacials) they have diminished to about their present extent with glaciers at sea level only near the poles. For the past 10,000 years glaciers have been generally in recession. However, from 100 to 40 million years ago it was much warmer in high latitudes and the pole-to-equator temperature difference was only 10 or 15°C, three times less than at present. There appears to have been no ice at sea level, and everywhere the mean surface temperature exceeded freezing (O°C). These are the characteristics of a "non-glacial period" and define Earth's normal climatic regime. Today, climatically speaking, we live in exceptional times. Figure 8(b) is a much simplified graph showing the variation with time of mean surface temperature in middle latitudes. During the early history of ericaceous plants, the Earth had an equable non-glacial climate (Fig. 8(b)). This was followed by a long slow decline to about 3 million years ago when the present glacial period commenced, although there is evidence of some glacial ice in Antarctica as long ago as 15 million years.
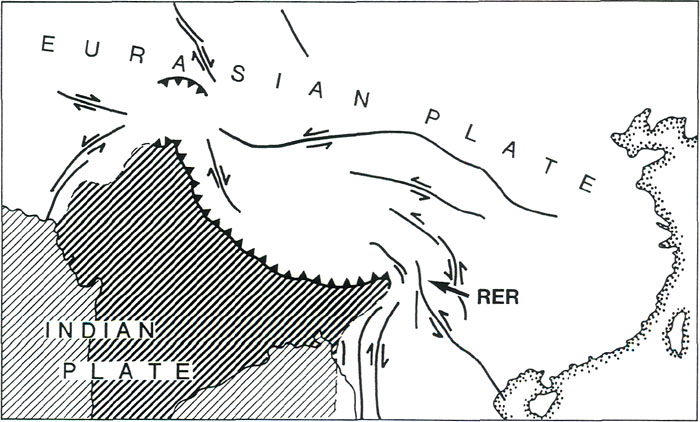
Figure 8. (a) Earth's mean temperature at sea level, compiled from Barron (1987). (b) Variation in mid-latitudes, of mean sea level temperature, compiled from Savin (1977) much simplified. Vegetation Zones For any configuration of global climate, the principle factor controlling climatic patterns is latitude. During the early history of rhododendrons 60 to 40 million years ago, North America and Eurasia were close to or north of the positions they occupy today; they were squarely lodged between 30 and 60°N latitude, with large areas north of 60°N (Figs. 5(a) and (b)). The climate, however, was warmer and more equable, and forest grew at much higher latitudes, because the Earth was then in its non- glacial state (Fig. 8). As a consequence, there appeared to have been only about four major vegetational zones, compared to about three times that number today. These are respectively from north to south, "tropical"; rain forest ("tropical"; placed in inverted commas because such forest extended well outside the tropics), rain forest, paratropical rain forest, broad-leaved evergreen forest, and polar deciduous forest (labeled P in Figs. 5(a) and (b)). These ancient forests can only be compared to modern forests in a general way because many modern forest trees had not yet evolved. For example, the dominant coniferous trees were not the spruces, pines and firs of today, but instead were, for the most part, members of the bald cypress family (Taxodiaceae), which includes such trees as the dawn redwoods, the redwoods and the bald cypress, whose range is nowadays fragmented and constricted. The polar deciduous forest, which included deciduous hardwoods and conifers such as the dawn redwood, occurred in northern North American and northern Siberia well inside the Arctic Circle. Around and just south of the Arctic Circle there was a belt of broad-leaved evergreen forest including palms, followed by a belt of warm paratropical rain forest. Both belts probably extended into lower latitudes in upland areas, and both probably provided habitats suitable for rhododendrons. "Tropical" rain forests were the dominant vegetation, which, apparently, covered most low-lying regions in latitudes lower than about 45°. Within it, most habitats suitable for rhododendrons are likely to have occurred only in upland regions, but these could have been widespread as already noted (Fig. 7). It seems probable therefore on geographic, climatic and ecological grounds that rhododendrons, during their early history, could have extended more or less continuously from North America to Greenland and Europe, and eastward into China and northeastern Asia. It is probable that they moved freely between North America and Europe across narrow gulfs of the nascent North Atlantic, rather then across Beringia, a route that was then at very high latitude. The early population of rhododendrons, although probably widespread, may not have been very diverse (that is, had comparatively few species). By 40 million years ago, there were about five global vegetational zones. "Tropical" rain forest occupied a smaller area than before (Fig. 5(b)). Polar deciduous forests persisted, and there was a notable development of a northern evergreen coniferous forest composed typically of members of the family Taxodiaceae. The belts of warm paratropical rain forest and broad-leaved evergreen forest moved southwards and expanded. Except for the region of extreme relief and the high-island archipelago which had not yet taken shape, the ranges of these two zones embrace the present-day distribution of rhododendrons, including that of the cold-climate R. lapponicum. It appears that, in and between the places where rhododendrons are found, rhododendrons are still more or less where they always have been. By 20 million years ago "tropical" rain forests were further restricted, and general cooling in middle and high latitudes had begun. These changes culminated in the glaciations of the past few million years. Continental climates and grasslands began to develop, and there was a further increase in the number of vegetational zones from about six at 20 million years ago, to as many as eight today, twice the number present when rhododendrons began (compare Figs. 5(a) and (d)). In the north, extensive coniferous forest dominated by the pine family (Pinaceae), came into existence. Its present-day descendant is the boreal forest of northern North America and northern Eurasia. Mixed hardwood and coniferous forest for the first time occupied large areas of Eurasia and North America, but the great broad-leaved evergreen forests of the early Tertiary were much diminished, becoming restricted as they are today to south-eastern China and the southeastern United States. These mixed hardwood and broad-leaved evergreen forests provided the habitats best suited to rhododendrons. Between 20 million years ago and today, and especially in the last few million years, the climate in middle and high latitudes becomes much cooler than when rhododendrons first made their appearance. Large areas of desert, semi-desert and grassland developed in central and southwestern North America, across Africa and into Asia (Fig. 5(d)). This expansion of arid habitats eliminated vast tracts of rhododendron habitat. During the past 2 to 3 million years, vegetational zones fluctuated greatly in response to glacial advances and retreats, which disrupted the climate of lands bordering glaciated areas. There was a broad circum-polar belt of tundra and ice, and "tropical" rain forests shrank to a remnant of their former extent. Today there are broad belts of desert and semi-deserts in tropical and mid-latitudes which are bordered by broad expanses of tropical and temperate grasslands, something quite unknown 50 million or more years ago when rhododendrons first appeared (Fig. 5(a)). The region of broad-leaved evergreens and mixed hardwood and coniferous forest is confined to eastern and southeastern North America, Europe and Asia. Extreme Relief and the Recent Speciation of Lepidote and Elepidote Rhododendrons One of the most spectacular events in Earth's history is the long drawn-out collision of India with Asia which began about 40 million years ago and continues today. This collision set in motion the train of events which lead to the creation of the region of extreme relief and, we argue, to conditions highly favorable to the development of a rich rhododendron flora. Prior to 120 million years ago, India formed part of the giant southern supercontinent, Gondwana, which comprised South America, Africa, Arabia, Antarctica and Australia, as well as India. Gondwana fragmented in piecemeal fashion. India and Australia were the last large fragments to break away. In its northward travel, India left behind an ever- widening rift which became the modern Indian Ocean. The ocean that once existed north of India slowly closed (Fig. 5). The Earth's crust beneath India is over two billion years old, and hence is cooler, denser and stronger than the much younger part of Asia with which it collided. As a consequence, India did not break and loose its identity, but relentlessly pushed northward, deforming much of southeastern Asia as it went. Immediately north of the zone of contact, the Earth's crust was doubled in thickness and was lifted up forming the Tibetan Plateau. South of the plateau, debris piled up forming the Himalayas (Fig. 6(a)). Together the Tibetan Plateau and the Himalayas form a vast region of high elevation unique on Earth (Fig. 3(a)). It is the only region where two continents of large mass are colliding with one another today. The question of the timing of uplift of the Tibetan Plateau and the exact manner in which it happened is a matter of much current debate. The most recent estimates are that rapid uplift began about 20 million years ago, and that the plateau reached approximately its present elevation about 8 million years ago (Harrison et al. 1992). The exact timing of uplift is important for understanding the details of the general vegetational history, and ultimately for understanding rhododendron evolution, but for our present purposes it is sufficient to know that there has been a region of high altitude there for at least 8 million years. The assembly of a landmass as large as Asia and the uplift of the Himalayas and Tibet has affected climate profoundly. Very low atmospheric pressure (centered over Pakistan and stretching northeast to Mongolia) develops in summer, drawing wet tropical air from across the equator, and causing heavy rainfall along the southeastern border of Tibet and the Himalayas (Fig. 3(d)). As uplift continued, river systems grew, their valleys etched along large crustal faults that had been developed as India pushed its way into Asia (Fig. 10). It is this close fan- shaped network of faults along the southeast border of the Tibetan Plateau that accounts for the geometry and bunching together of the deep valleys. The fault-controlled alignment of valleys provides ready access to rain-bearing prevailing winds (Fig. 3) ensuring that the slopes of the region of extreme relief are well watered. Through time, a landscape evolved which became home to a wide variety of habitats - sub-tropical valley bottoms, temperate slopes, and alpine peaks all very close together.
Figure 9. Maximum of ice sheets during glacial period of past few million years.
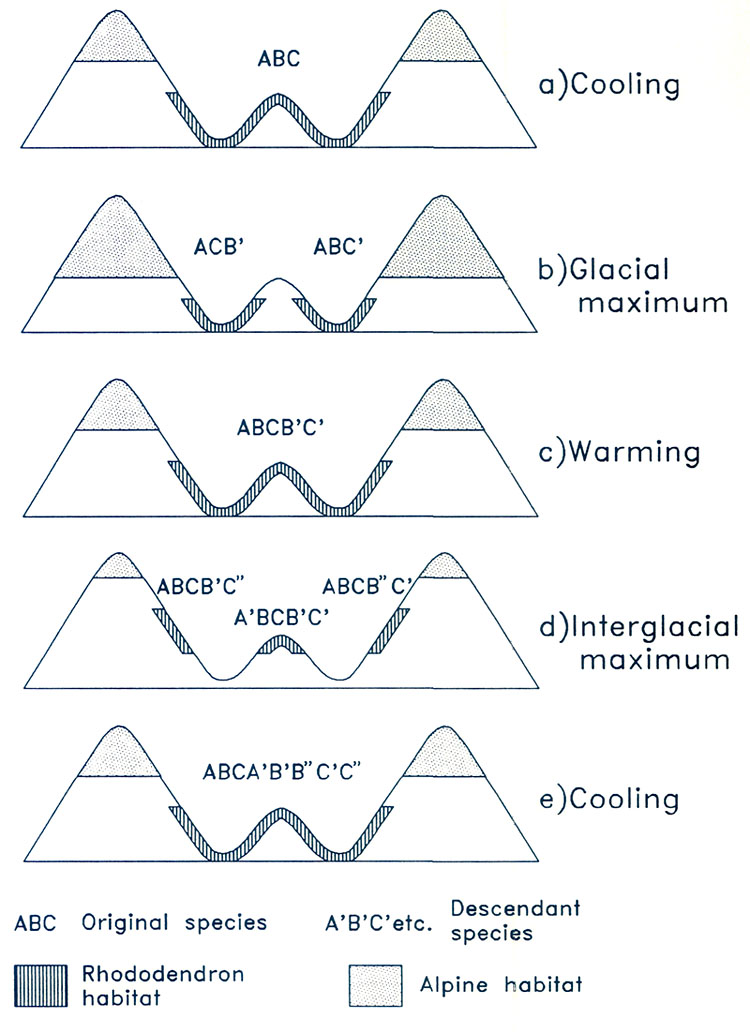
Figure 10. Great shear faults which define the river valleys in the region of extreme relief (RER). Compiled from Molnar and Taponnier (1975). The assembly of a landmass as large as Asia and the uplift of the Himalayas and Tibet has affected climate profoundly. Very low atmospheric pressure (centered over Pakistan and stretching northeast to Mongolia) develops in summer, drawing wet tropical air from across the equator, and causing heavy rainfall along the southeastern border of Tibet and the Himalayas (Fig. 3(d)). As uplift continued, river systems grew, their valleys etched along large crustal faults that had been developed as India pushed its way into Asia (Fig. 10). It is this close fan- shaped network of faults along the southeast border of the Tibetan Plateau that accounts for the geometry and bunching together of the deep valleys. The fault-controlled alignment of valleys provides ready access to rain-bearing prevailing winds (Fig. 3) ensuring that the slopes of the region of extreme relief are well watered. Through time, a landscape evolved which became home to a wide variety of habitats - sub-tropical valley bottoms, temperate slopes, and alpine peaks all very close together. Speciation occurs more readily in small reproductively isolated populations of plants and animals than in large contiguous populations; favorable genetic modifications spread much more rapidly through small compact populations than through large diffuse ones distributed over a wide area; in a small population, genetic drift is more likely to cause changes in a population’s characteristics. Such small reproductively isolated populations that undergo rapid diversification are referred to as "founder populations" (Mayr 1982). What were the founder populations of the present-day, highly speciated rhododendrons of the region of high relief, where were they located, and where did they come from? As already explained, it is possible that during the period from about 60 to 40 million years ago, rhododendrons ranged more or less continuously across North America and Eurasia because suitable climates and topographies occurred there over a much wider area than at present (Figs. 5(a) and (b), and 7). Although rhododendrons may have been widespread and abundant, the uniformity of conditions may not have lead to the evolution of many species. As Asia was enlarged by the addition of India, and as the Tibet-Himalaya region was uplifted, new habitats ideal for rhododendrons came into being. It is possible that representatives of both lepidote (subgenus Rhododendron) and elepidote (subgenus Hymenanthes) rhododendrons, but not azaleas, expanded into these newly established favorable habitats and became founder populations. Exactly where they came from cannot be determined in the absence of fossil records, but the mixed hardwood, coniferous and broad-leaved evergreen forests of eastern Asia about 20 million years ago are their most likely source (Fig. 5(c)). It is of vital significance for rhododendron evolution that the rapid uplift of the Tibetan Plateau was occurring at a time when global climate was cooling. As Earth entered a glacial period, ice advances and retreats occurred repeatedly every 50 to 100 thousand years. In the region of extreme relief, as an ice age commenced and harsh conditions at high altitudes expanded, species ranges would be displaced downward to lower elevations (Fig. 11). Populations in valley bottoms, previously separated by unfavorable condition, could mix with populations from adjacent slopes and valleys, and species could cross after long intervals of separation to generate new hybrids, the potential progenitors of new species. Populations at higher elevations would become separated temporarily from one another by intervening glaciers or unfavorable alpine habitat. These now-separated populations would reproduce in isolation and, over time, diverge from each other by genetic drift following their own evolutionary path. During interglacial intervals, populations at higher elevation could migrate upward over ridges and divides, and formerly isolated populations would intermingle. The large vertical range of valleys and ranges meant that species distributions could be displaced rapidly upward and downward without annihilation, despite major climate shifts, because the actual distances traveled would be short. Consequently, fragmentation and coalescing of rhododendron habitats could have occurred repeatedly as a consequence of glaciation and deglaciation, so producing the present-day diverse flora.
Figure 11. Hypothesis of speciation during glacial cycling in region of extreme relief. (a) During cool period following an interglacial interval, species A, B and C are distributed across lower valley slopes and the central lower divides. (b) During a glacial maximum, the species disperse down slope becoming separated by the central divide, and B species in one valley and C in the other change to B' and C' through genetic drift. (c) Warming allows population to ascend, and five species are now everywhere present. (d) Warming continues so that this mixed population disperses upward and is split into three; genetic drift produces three new species, C' from C on the left-hand slope, A' from A on the central ridge, and B' from B on the right-hand slope. (e) Cooling causes populations to descend and remix. During the cycle, the number of species has been increased from three to eight. The whole cycle is assumed to take between 50,000 and 100,000 years. In essence, mountains and deep valleys each act as islands of habitat which repeatedly join and separate as climatic zones and plant populations ascend and descend. These joining- separating cycles occur at about the right interval for them to serve as a forcing process for speciation. A period of 50 to 100 thousand years is generally considered sufficient for a small, physically isolated population to undergo changes great enough to ensure reproductive isolation. These separations and rejoining would provide repeated opportunities for species to mix and separate repeatedly. The end result would be a very large number of closely related species many of which would retain the ability to hybridize. We suggest that it was precisely this combination of very high mountains situated in tropical latitudes, and cycles of glaciation and deglaciation, that provided the dynamic conditions for the extraordinary speciation of lepidote and elepidote rhododendrons in the region of extreme relief. According to this view, most of the speciation has happened in the last 3 million years during the current glacial period. Lowland areas or mountains of modest size provide fewer opportunities for populations to separate and then rejoin (the hallmark of the above speciation mechanism). They provide no refuge in times of severe climate. In valleys of less than a certain critical depth, rhododendron populations would be eliminated or greatly reduced as climate deteriorated; as temperature fell and valleys become occupied by unfavorable habitats even filled with ice, there would be no possibility of downward migration; as rainfall diminished during warmer interglacial periods, there would be no nearby higher wetter slopes on which to find refuge. Critical valley depth depends mainly on latitude. Western North America or the Alps, for example, have about one half the relief and are more northerly than the region of extreme relief. Much of northern North America was covered by ice during major glaciations, whereas to the south relatively dry climate prevailed. There were few suitable habitats nearby, and during ice ages the local rhododendron population must have suffered severely. In the region of extreme relief the correct balance appears to have been struck, so that although climatic changes were great, the tropical latitudes and the great depth of the valleys ensured that safe havens were nearby. In such a physical environment, latitudinally well-placed, climate change acts not as an agent of destruction, but as an incentive to diversification. Nowadays, because of its unique elevation, the region of extreme relief is isolated botanically from the rest of the Asian land mass of which it is a part. Moreover, the region itself is a vast intricate honeycomb of valleys whose degree of isolation changes as ice ages wax and wane and as continuing uplift and erosion modify their configurations. Figure 12 depicts the process of evolution proposed above. Azaleas Azaleas differ from other rhododendrons in that there are only about one tenth the number of species, and they are, so far as we have been able to determine, absent from the region of extreme relief. Deciduous azaleas occur mainly in southeastern USA and in south-eastern Asia and Japan, with scattered representatives in southern Europe, Asia Minor, southwestern Asia, and western North America. This pattern indicates that, as has happened with the subgenera Hymenanthes and Rhododendron, they were more generally distributed over the northern hemisphere, and that their range has since been reduced and fragmented by climatic deterioration and by the widening of the North Atlantic, which increasingly presented a barrier to dispersal. Evergreen azaleas are found only in southeastern China and Japan, and it is possible that they have always remained confined to warm temperate broad-leaved evergreen forests. They may never have ventured far beyond their original ecological niches, and their range has, in consequence, shrunk along with that of broad-leaved evergreen forests (Fig. 5). The azaleas are not plants of the region of extreme relief although they live close by in terrain of about one half or one third the altitude and relief. Presumably, because of their genetic make-up and historical circumstances, they were unable to take advantage of the dynamic environment of the region of extreme relief. Although well positioned to act as founder populations, they failed to do so. In this respect, the paleogeographical evolution of azaleas appears therefore to have been very different from that of other rhododendrons (Fig. 12).
Figure 12. Proposed paleogeographic evolution of rhododendrons. Except for the evergreen azaleas (Tsutsusi) which may always have been fairly restricted, rhododendrons were, we suggest, spread widely across the northern continents during the climatically mild interval 60 to 40 million years ago. Although widespread and abundant, they were not necessarily represented by many species (not strongly diversified). As climate in middle and high latitudes began to deteriorate about 20 million years ago, and as the North Atlantic opened, suitable habitats were restricted and rhododendrons became confined to isolated pockets. At about this time, we suggest that small founder populations (circled in the diagrams) of the subgenera Rhododendron (lepidote) and Hymenanthes (elepidote) colonized what is now the region of extreme relief as Tibet began to be uplifted, and the deep valleys on its southeastern border were eroded. These founder populations proceeded to speciate, especially during the past few million years. The vireyas, an offshoot from the region of extreme relief, were a late development which colonized the recently formed high-island archipelago. High-island Archipelago and the Speciation of Vireyas The Indo-Malaysian region has an abundance of species from section Vireya of the subgenus Rhododendron. Other representatives of the genus are rare (the evergreen azalea R. subsessile grows in the Philippine Islands). Vireyas are also found, but in much fewer numbers, in the region of extreme relief, in the mountain ranges of Indochina and Malaya, and in adjacent areas of southeastern China and Taiwan (Fig. 1 d)). In contrast to many other sections, such as the section Ponticum (of the sub- genus Hymenanthes) which has representatives in North America, Europe, and southeastern Asia and a markedly discontinuous distribution, the vireyas are concentrated in one essentially continuous region. This suggests that they have originated comparatively recently, more recently, for example, than the section Ponticum. The terrains in which the majority of vireyas are found today are of recent origin indicating that the abundance and diversity of vireyas is also a recent phenomenon. Many of the mountain ranges of the high- island archipelago, have been created by volcanic activity as a result of recent subduction and melting of the Indian and Pacific Plates (Figs. 3(b) and 6(b)). In their current manifestations, they are probably of recent origin, no more than a few million years old. A case in point is the highlands of New Guinea, which exceed 4000 meters in relief and which began to be uplifted about 4 million years ago (Hill and Gleadow 1989). Because the region is near the equator, vegetational zones vary from tropical forest at sea level to alpine grassland with glaciers on the peaks, and they may have fallen as much as 1000 meters during ice ages (Hope 1986). Hence, a mechanism for speciation similar to that proposed for the region of extreme relief may also be operative here. We suggest that the founder populations of the section Vireya originated in the region of extreme relief within the past few million years. From there they have spread swiftly down the Malay Peninsula into the high-island archipelago where they speciated rapidly (Fig. 12(b)). One species has even reached Australia, where it occurs in the tropical rain forest of the Atherton Tableland. Summary The essence of our thesis is that, following a period of mild climate when rhododendrons were widely and more-or-less continuously distributed across North America and Eurasia, their range became much reduced as a result of global climatic deterioration. This deterioration began about 25 million years ago, was clearly marked by about 15 million years ago, and became extreme with the onset of the current glacial period about 3 million years ago. Small founder populations of the subgenera Hymenanthes and Rhododendron (but not azaleas) situated marginal to their main range were able to enter and retain a foothold in the newly developing region of extreme relief on the southeastern fringe of the Tibet-Himalayan region. There they proliferated, and it was from there that the vireyas spread into the mountains of the high-island archipelago. By taking advantage of special newly developing conditions, these small, originally peripheral populations have become now the most numerous and diverse. The place of origin of rhododendrons is not known, but is was not the region of extreme relief where they are nowadays most diverse and abundant. Many of the arguments we have presented are very speculative, necessarily so because of the poor representation of rhododendrons in the fossil record. Nevertheless these arguments provide reasonable explanations of their present remarkable distribution and of their differing abilities to cross and recross. Acknowledgements We are very grateful to Darlene Upton and Lara Pearson for preparing the manuscript, to Gina Lesperance and Linda Bedard for preparing the figures, all of the Pacific Geoscience Centre; to Dr. Glen Rouse of the University of British Columbia for directing us to papers on the fossil record of rhododendrons and vegetation history; to Mr. Norman Todd and Dr. Alec McCarter of Victoria, to Dr, Geraldine Allen of the University of Victoria to Mr. Frank Doleshy of Edmonds, Washington, and to Dr. John Fyles of The Geological Survey of Canada for constructive criticisms of the manuscript. References
1. Barron, EJ., Paleoceanography, 2,729-739,1987. Ted Irving, a retired geophysicist, is an emeritus scientist with the Geological Survey of Canada, Richard Hebda, a botanist with the Royal British Columbia Museum. Both are adjunct professors at the School of Earth and Ocean Sciences at the University of Victoria. Editor's Note: Other authors have discussed the origin and distribution of the genus Rhododendron in articles in the Journal American Rhododendron Society, most notably Frank Doleshy ("Distribution and Classification of Certain Japanese Rhododendrons," J. Amer. Rhodo. Soc. 37:81-89, 1983, and "How the Japanese Pontica Rhododendrons Fit Together," J. Amer. Rhodo. Soc. 42:190-194, 234- 235, 1988). Three other Journal authors who have explored the subject are W.R. Philipson and M.N. Philipson ("Rhododendron Emigrants," J. Amer. Rhodo. Soc. 39:29-30, 1985 and L. K. Wade, "Mount Kinabalu - A Botanist's View," Amer. Rhod. Soc. 35:62-73, 1981.) Numerous field observations of rhododendrons growing in the wild today have been made and discussed in Journal articles. Among these authors are Peter Cox, Warren Berg, Gwen Bell, Dallas Boge, Michael McCullough, Frank Mossman, Britt Smith, Pieter van Royen, Clive Justice, Dave Goheen, Bill Jenkins, Udai Pradhan, Bengt Skoog, Mark Widrlechner and many more.
|
|||
|
|
|||